Nerve Muscle Physiology
Feb 19, 2025
Navigate Quickly
Nerve
Blood-Brain Barrier
Structure of a Single Neuron
Functional Areas of the Neuron
Different Types of Neuron
Axonal Transportation
Resting Membrane Potential
Arrangement of Resting Membrane Potential
Origin of Resting Membrane Potential
Leak Channels
Nernst Equation
Action Potential in Neuron
Action Potential
Different Parts of Action Potential
Permeability of IONS During Action Potential
Potassium Channel
Refractory Period
Different Stages of Voltage-Gated Sodium Channel
Nerve Fibre Classification
Classification of Nerve Fibres ;
The function of the Fibres
The Conduction Velocity of the Nerve Fibres
Compound Action Potential
Stimulus Parameters
Utilization time
Chronaxie
Excitability of the Tissue
Strength-Duration Curve
Injury to Nerve Fibers
Sunderland’s Classification
Seddon’s classification
Sarcomere
Structure of the Sarcomere
Length of the Sarcomere
Starling’s Law
Muscle proteins
1. Dystrophin
2. Titin
Important Points to Remember
Important Solved Questions


Nerve
There are multiple bundles of nerve fiber in the cross-section of any peripheral nerve of the body. These bundles of nerve fibers are called fascicles. Inside this fascicle, there are multiple nerve fibers; these are the axons of a single neuron. The nerve fiber is surrounded by a connective tissue layer. This connective tissue covering is known as endoneurium.
The whole fascicle is also surrounded by connective tissue called the perineurium. The nerve fibers are surrounded by a loose connective tissue covering called the epineurium. The capillaries that supply blood to the neuron are located in between the fascicles. Perineurium forms the blood neuron barrier as it consists of connective tissue cells and between these connective tissue cells there is a minimum gap. These gaps are bridged by tight junctions (composed of occludin and claudin). So, the perineurium and the tight junction form a barrier called the blood neuron barrier. (As per research articles not only Perineurium but even the endothelium cells of the capillary are also bridged by tight junctions).
Read this blog further to get a quick overview of this important topic Nerve Muscle Physiology and ace your NEET PG exam preparation.
Blood-Brain Barrier
The brain capillaries are also lined with endothelium cells. The capillaries present in the brain are continuous in nature. Continuous capillaries mean that the gap between the capillaries is very less, which is about <2μm. This gap is bridged by tight junctions. Astrocytes are not the part of the blood-brain barrier; it only regulates the permeability of the blood-brain barrier.

Structure of a Single Neuron
- Dendrites are known as afferents because all the signals are received here.
- Soma is the cell body of the neuron, also known as perikaryon. The size of a neuron is variable.
- The smallest cell body is present in the granule cell of the cerebellum 5μm.
- The largest cell body is present in the neuron of the Anterior horn of the cerebellum 135μm.
- Nissl body is the aggregate rough endoplasmic reticulum and some free ribosomes.
- They are very rich in RNA material because of which the stained Nissl body will be basophilic in nature.
- It is never present in the axonal cytoplasm.
- The highest density is found in the motor neuron which is metabolically very active.
- Cell cytoskeleton
- The main cytoskeleton organelle is an intermediate filament which is called neurofilament, which is responsible for cell size.
- Microtubules and microfilaments are also present inside the cytoskeleton.
- Centrioles
- They are also present in adult neurons as well.
- Centriole maintains the microtubules inside the cell cytoskeleton.
- Pigments
- Melanin pigment is present at Substantia nigra neurons, which is why it is brown in color. It is also present in the Locus coeruleus nucleus, resulting in a blue color.
- Iron (Fe) at the red nucleus.
- Zinc (Zn) at the hippocampus.
- Axon
- Axon Hillock: This is a part of the cell body where the membrane is thicker and is also attached to the axon. The first of the Axons are about 50-100μm which starts after the axon hillocks, this is known as the initial segment. Axons at the terminal level will be divided into multiple small vesicular regions which are known as Axon Knob. Myelin Sheath in a long neuron can be 1mm in length. In between two myelin sheaths, the region is 1μm in length and this is known as the Nodes of Ranvier.
Also read: Body Fluid Compartments and its Measurement
Functional Areas of the Neuron
Functional Areas
- Graded Electrogenesis → Dendrites & Soma
- AP initiation
- Voltage gated Na+- channel
- Max
- NOR (2000 – 12000/μm2)
- Initial segm. (350 – 500)
- Soma/dendrites (50-75/μm2)
- Telodendria (20-75/ μm2)
- Least → On surface of myelin sheath
Different Types of Neuron
Unipolar Neuron: A unipolar neuron only has one nerve process extending from the cell body.According to the genome Unipolar neuron is only present in invertebrates. But according to the anatomy book it is given that the unipolar neuron present in our body is the mesencephalic nucleus of the fifth cranial nerve.
Pseudo Unipolar Neuron: A neuron that develops as a bipolar neuron but eventually merges to form a unipolar neuron. Example: Sensory neuron.
Bipolar Neuron: It is a type of neuron that has two extensions (one axon and one dendrite). Bipolar neurons are located at the level of the retina. Example: Retinal bipolar cell and olfactory receptor neurons.
Multipolar Neuron: A multipolar neuron is a type of neuron that possesses a single axon and many dendrites. Example: Motor Neuron.
Axonal Transportation
Nissel's body produces a protein molecule, for instance, this protein molecule may be a channel etc. The transportation of the protein from the level of the cell body to the axon end is nothing but axonal transport/ intracellular transport. This process is similar in every cell, but the process is well-studied in the case of neurons. In the case of the neuron the transportation within the cytoplasm is known as axonal transport.
Process- The protein will be packed inside the vesicles; this packaging is done by the Golgi bodies. Inside the axon there are certain cytoskeletons; one of the important cytoskeletons present is microtubules. (Microtubules are ever-changing, with reactions constantly adding and subtracting tubulin dimers at both ends of the filament. one end grows more rapidly and is called the plus end, whereas the other end is known as the minus end). Microtubule is the pathway for the transport of the vesicle. On this path there are molecular motor systems running on this pathway which are known as Kinesin. (It is a plus end-directed motor). Then this kinesin will move until it reaches the end of the microtubule and it will release the vesicle within which the protein is present. Opposite transportation from axonal end to cell body this type of transportation is a retrograde transport.
- Molecular Motor
- Kinesin + end directed transport.
- Dynein - end-directed transport.
- Myosin (myosin type 5, 1 and 6) bi-directional transport.
- Types of Axonal transport as per velocity
- Fast anterograde: Molecular motor kinesin, Speed400 mm/day, it transports vesicles and mitochondria.
- Slow retrograde: Molecular motor dynein, Speed 200mm/day, transports viruses, toxins and growth factors.
- Slow anterograde: Molecular motor not known, Speed 0.5 to 10mm/day, it transports components of microtubules and microfilament.
Resting Membrane Potential
Resting means when the cell does not receive any external stimulation or invasion. In the resting case, most of the cell remains in equilibrium condition. Potential refers to charge of the cell. The membrane voltage difference between inside the cell and outside is known as resting membrane potential. Conventionally, outside cell reference is electron reference point (ECF) and interior cell reference (ICF) is called active or recording electron. The voltage difference of inside with respect to outside is known as resting potential difference (RMP). Different cells have different resting potentials. But in most of the cells, RMP ranges from -10 mv to -100 mv. To measure the resting potential difference of the cell, one electron is introduced to the inside and one outside cell membrane. A voltmeter is connected between them to measure the potential difference. This newly added electron must be very thin or it will rapture the cell A glass microelectrode filled with KCL solution with tip diameter of 0.5 μm is used for recording RMP.
Also read: Chemical Regulation of Respiration: Role of Chemoreceptors and Reflexes
Arrangement of Resting Membrane Potential
An electrode filled with KCL solution is placed just beneath the cell membrane. This electrode is known as microelectrode which is very thin with a tip diameter of 0.5 μm. Metals cannot be used for this type of electrode so the electrode used here is made up of glass material. A glass electrode filled with KCL solution is inserted into the cell. This electrode has a recording wire which is connected to a sensitive voltmeter. The other end of the voltmeter is connected to an electrode that is placed outside the cell membrane. Thus with the help of a glass electrode and very sensitive voltmeter, we can easily calculate the resting potential difference between the two electrons. Cathode ray oscilloscope is used to record a graphical image of resting potential. It is a computerised recording screen that can capture the graphics of resting membrane potential.
Origin of Resting Membrane Potential
Membrane potentials caused by concentration differences of ions across a selectively permeable membrane. Uneven distribution of ions between ECF & ICF.Different permeability to different types of ions in resting stage. Contribution of the potassium diffusion potential. Contribution of the Na+ - K+ Pump. Membrane potential is caused by concentration difference of ions of a selective ion across a selectively permeable membrane.
Hence two major requirements for generation of RMP are:Unequal distribution of ions between ECF and ICF and Membrane should be selectively permeable to different ions.
Donnan’s effect plays an effective role in creating and maintaining uneven distribution of ions between ECF and ICF. The channels that are open even in the resting condition of the cell are known as leak channels. They are always open and diffusion continues to occur through them. Leak channels are responsible for the generation of resting potential of the cell. Potassium is the main ion that diffuses through the leak channel and generates resting potential difference. The NA-K-ATPase pump also plays a vital role in generating RMP. As the pump throws three positively charged molecules of sodium outside and takes only two positively charged molecules of potassium inside it. This automatically turns the membrane voltage to a negative side. Thus NA-K-ATPase pumps keep the RMP negative. For example if the RMP of the cell is -65 nm. If a NA-K-ATPase pump is added to it RMP will become more negative. In general NA-K-ATPase pumps contribute 5 to 10% of RMP generation.
Donnan Equation
A semipermeable membrane divides the box into two compartments A and B that are filled with solution. When sodium ions are injected into these compartments assuming that the membrane is permeable to sodium and chloride, When 9 molecules of positively charged sodium ions are infused on both sides, no diffusion will take place as they are equal in composition. Now, if 9 molecules of chloride are infused on side B, it will try to shift on side A. After a certain point of time equilibrium will be created. This means that 4 molecules of chloride will be present on side A and 4 molecules of chloride will be present on side B. The extra chloride molecule will keep on transferring from side A to B. At this point of time, if we add 9 large negatively charged impermeable protein molecules on side A, all the dynamics will change. Although the negatively charged protein cannot travel, it will attract all the positive charge from side B and repel all the negative charges.
Thus attraction of sodium and repulsion of chloride molecules will continue for a certain period of time after which equilibrium state is established. The next example represents the equilibrium state wherein sodium molecule inside B will be less (say 6) and it will be more inside A (say 12) and the number of chloride molecules will be 6 inside B and 3 inside A. Usually the chloride molecule is distributed equally between both sides but due to repulsion force exerted by protein, there’s a decrease in chloride shift and increase in sodium molecule from side B to A. Also nine molecules of negatively charged protein are present inside A. This is the equilibrium state. This condition was first described by Donnan and therefore is known as Donnan’s effect. According to Donnan, due to the presence of a large impermeable molecule, distribution of permeable ions or molecules changes through a semipermeable membrane.
Donnan revised this phenomenon along with Gibbs to introduce a few mathematical equations. According to them, the total amount of positive and negative charges in compartment A (12) is equal to the number of negative charges in compartment A. [T+]A = [T-] A Therefore it can be concluded that in equilibrium state, compartment A is an electro neutral compartment where total positive charge is equal to the total negative charge. Total positive charge of compartment B is also equal to the total negative charge of compartment B [T+] B = [T-] B
Therefore, compartment B is also an electro neutral compartment where total positive charge is equal to the total negative charge. When we consider the permeable ions only and neglect protein, then sodium multiplied by chloride ion on compartment A is equal to the sodium multiplied by chloride ion on compartment B.[Na×Cl] A = [Na×Cl] B Product of the permeable ion concentration on compartment A (12×3=36) is equal to the product of permeable ion concentration on compartment B (6×6=36). This equation is only applied to permeable ions. Due to the equilibrium state both sides become electro neutral. However, the total number of ions inside A is high (12+3+9) and but total number of ions in side B is less. As a result of the large number of ions present inside A, the osmotic tension of this compartment will be a lot more than that of side B.
Also read: Cell Physiology: Overview of Membrane, Cytoskeleton & Junctions
Leak Channels
Certain channels are open on cell membranes which are known as leak channels. In the above diagram a resting cell is shown with some leak channels
In most of the mammalian cells, potassium sodium and chloride leak channels are present which are always open.
The simplest form of leak channel any cell can have is potassium leak channel.
Glial cells contain only one type of leak channel that is potassium leak channel. Since it is a mammalian cell, the potassium concentration inside the cell is higher than that of outside membrane.
As we know that the cell is an electroneutral compartment which has equal number of negative and positive charges, therefore potassium also contains equal amounts of negative charge which may or may not be permeable through the cell membrane. Similarly the outside potassium is also accompanied by corresponding negative charges. Due to the presence of chemical gradient or concentration difference between inside and outside the cell, potassium will try to shift outside the cell through the leak channel and since the potassium is coupled with a corresponding negative charge, the negative charge also moves along with the potassium to the leak channel.
When charge of the coupled ions reach potassium leak channel gate, it will only allow potassium to pass and the negative charge is withheld from transferring and it will get deposited at the surface.
As more potassium passes through the leak channel, a number of negative charges get deposited at the inner leaflet of a cell membrane.
At this point the cell membrane has extra positive charges on the outer side and extra negative charge on the inside. Thus due to potassium diffusion, the inner and outer leaflet of the membrane have different numbers of charges.
Nernst Equation
According to Nernst equation, equilibrium potential of an ion is equal to RT/ZF log of concentration of ion outside the cell upon concentration of the same ion inside the cell.
Eq = RT/ZF.log [x]outside/[x]inside
This equation is given in natural logarithm. To convert it into log base 10, we have to multiply it by 2.3. Thus the equation becomes:
Eq = 2.3 × RT/ZF.log10.[x]outside/[x]inside
Where RT/F is gas constant having value of 23.7 mv at 37° C
[x] is the concentration of ion inside and outside and
Z is the volume of molecules used.
This constant changes with the change in temperature. And therefore the value of this constant varies in different textbooks as most of the experiment is done on seawater whose temperature is different from our body temperature.
Upon multiplying 2.3 to gas constant, the equation becomes:
Eq = +61/z.log10. [x]outside/[x]inside
This is the simplified version of Nernst equation.
EQUILIBRIUM POTENTIAL OF IONS IN VARIOUS MAMMALIAN CELLS
| IONS | EP (Acc to GYTON) in mV | EP (Acc to Others) in mV |
|---|---|---|
| Sodium | +63 | +65 |
| Chloride | -71 | -75 |
| Potassium | -90 | -94 |
| Calcium | +132 | +132 |
All the above values are calculated using Nernst equation with temperature of 37° C that is normal human body temperature. Almost every mammalian cell is permeable to three different ions that are potassium, chloride and sodium molecules. All of these leak channels will try to attain equilibrium conditions. In such cases RMP will be in constant motion. As the number of leak channels and permeability of potassium is much higher than of chloride and sodium, therefore RMP will finally move to potassium. This is known as the permeability coefficient of these ions.
The permeability coefficient of potassium, chloride and sodium is 1:0.45:0.04. This means that in a mammalian cell, if there are 100 potassium channels open, there will only be 45 Chloride and 4 sodium League channels open in that cell.As RMP moves close to equilibrium potential of potassium, we can conclude that in every typical mammalian cell, RMP is close to equilibrium potential of potassium which is the maximum permeable ion.
In any vertebrae cell, muscle or neuron cells, RMP is generated by all the various ions present in that cell but it will move close to the maximum permeable ion present.
Thus we can conclude that if the cell is permeable to only one type of ion then RMP will be equal to the equilibrium potential of that ion. This RMP can be calculated by using Nernst equation. But if the membrane is permeable to three different ions then every ion will contribute in generating the RMP but RMP will move closer to the maximum permeable ion.
Above cited equation can be used to calculate the RMP of a cell membrane which is permeable to more than one ion.
- Where, Vm is the membrane voltage,
- RT/F is the gas constant, in natural logarithm.
- Pk is the permeability of potassium,
- Pcl is permeability of Chloride,
- pNa is permeability of Sodium.
- K+, Cl-, Na+ are the ion concentration.
- I stands for inside concentration and O is the outside concentration.
This equation is known as Goldman-Hodgkin-Katz equation or simply Goldman constant field equation.
Also read: Biostatistics Multiple-Choice Questions for Health Sciences
Action Potential in Neuron
Action Potential
When an excitatory neuron gives stimulation to the neuronal body, the postsynaptic membrane gets the stimulation. -70 is the RMP. The membrane voltage will change gradually towards depolarization. Whenever the membrane voltage is crossing - 55 mv, it is very important because it is the threshold voltage of the neuronal membrane. Whenever the membrane voltage is crossing the threshold voltage, the voltage gated Na+ channels present at the area of the initial segment will open up simultaneously leading to a huge amount of Na+ entering the cell causing the membrane voltage to rise to +35mV. This is called Action potential generation of membrane.An action potential is a rapid sequence of changes in the voltage across a membrane. The membrane voltage, or potential, is determined at any time by the relative ratio of ions, extracellular to intracellular, and the permeability of each ion. When the membrane voltage is rising from -65 to -55mV, this is called depolarization. This depolarization is known as Local potential or Graded potential.
- The properties of Action potential
- Non-propagated
- Amplitude of the local potential will depend on the strength of the stimulus in a graded fashion.
- The local potential is a Decremental potential.
- Local potential does not follow “All or none law”.
- The properties of Action potential will be opposite to the local potential i.e.,
- Action potential is a propagated potential.
- It is a non-graded potential.
- It is a non-decremental potential.
- The action potential will follow All or none law
- Action potential does not have summation properties.
- Summation of local potential.
- Adding up two local potentials is called summation of local potential.
- An excitatory neuron is producing +10mV local potential in the neuronal body, if the stimulation is stopped the local potential will die out at the same area by leakage of ions from the cell body. The local potential will move a very short distance unlike the action potential which is a propagated potential.
- LENGTH CONSTANT: The distance at which the local potential is reduced to 37% of the original voltage is known as the length constant. It is denoted by Lambda (Λ/λ). Generally, in the neuron the Lambda (Λ/λ) = 1-2 mm.
- TIME CONSTANT: When the local potential is not travelling at all, then also it can reduce to its original voltage over time. So, the time constant is nothing but denoted by τ (tau), the time at which the local potential reduces to 37% of the original.
- Temporal summation: Temporal summation means that the effects of impulses received at the same place can add up if the impulses are received in close temporal succession.
- Neurons are given two stimulation in succession. The first stimulation of the local voltage generated here is +5mv, which will decrease over time but before this decrease, if the neuron receives the second stimulation of +5mv. So, these two simulations will sum up together only in the case of a long time constant, and for instance, if the threshold voltage of the neuron is 10 mv then obviously, this local potential will be able to produce an action potential.
- Therefore, if there are two stimuli given in the same area to the neuron in order to sum them up, together then a long-time constant is needed. This kind of summation is called temporal summation.
- Spatial summation: Spatial summation occurs when stimuli are applied simultaneously, but in different areas, with a cumulative effect upon membrane potential.
- For spatial summation, two neurons are required to give simultaneous simulations. The local potential of these neurons will move towards the initial segment. If their length constant is long enough, these potentials will reach the initial segment and add up with each other. If the length constant is short, the local potential will move towards the initial segment but won’t be able to add up with each other. Therefore, the Spatial summation occurs when stimuli are applied simultaneously, but in different areas, with a cumulative effect on membrane potential. Spatial summation uses multiple synapses acting simultaneously.
- Whenever a summation takes place, the ultimate aim is to cross the threshold voltage, after crossing the threshold the target is opening the voltage-gated sodium channels. Opening these sodium channels will lead to huge amounts of sodium entering the cell which will raise the membrane voltage to +35 mv which is nothing but the action potential.
- If one electrode is connected inside the cell and one is connected outside the cell with a screen to record (CRO), we will get a graph of action potential on the CRO.
Pharmacology Related Articles:
Different Parts of Action Potential
- Whenever there is stimulation on the neuron. There will be a gradual increase in the membrane voltage towards the threshold potential and this is called local potential. This is due to the entry of sodium through a few sodium-gated channels opening up.
- Whenever the local potential is touching the threshold a positive feedback phenomenon will occur, the local potential has already developed some depolarization on the membrane and if the depolarization is sufficient enough to open the sodium channels. Further sodium will enter which will cause further depolarisations. This phenomenon is called the positive feedback phenomenon. There will be a rapid rise of membrane voltage to +35 mv due to rapid opening of Na+ gated voltage channels with a fraction of a second which is called the depolarization phase of an action potential. Time required to open this channel is 0.1 millisecond and the time required for inactivation is 1 millisecond.
- Whenever there is inactivation of the sodium channel the membrane voltage will not rise further. But proportionally the potassium channels will also open up, which means during the depolarization phase sodium is entering and the sodium channel is inactive but the potassium channel is open resulting in the potassium flowing out. If the potassium is going out the cell is losing a positive charge and the membrane voltage will be more negative. This is called the repolarization phase of the action potential.
- The voltage-gated potassium is very slow, which leads to the hyperpolarization phase of the action potential. This is caused due to the potassium gate being slow, when the membrane voltage is returning towards the RMP. Ideally, when the voltage is at RMP all the Potassium channels should be closed. But because of the slowness of the potassium channel, although the membrane voltage has reached the RMP, all the potassium channels are not closed yet. So, if the potassium channels remain still open some extra potassium will go out of the cell which was not supposed to happen. This loss of potassium from the cell will result in more negative voltage. This rise of negative voltage is known as the After hyperpolarization phase of an action potential.
Permeability of IONS During Action Potential
Sodium Channel
Never there is a generation of local potential the sodium channel will start opening. In the above diagram, the green colour curve shows the sodium channel opening. When it reaches the threshold voltage the number of sodium channel openings will rise as shown in the above graph. After reaching the maximum height the sodium channel will become inactive. This green colour curve is called the gNa+ (g- conductance which means permeability). The green colour curve shows how many sodium channels are open during the action potential.
Also read: Neural Regulation of Respiration: Control & Mechanisms
Potassium Channel
The potassium channel will also start opening with the sodium channel but since the sodium channel's permeability curve will appear different. The permeability of the potassium channel during the action potential is shown with the purple colour curve in the above graph. The opening of K+ channels will take a long time and closing also will take a long time. This curve is called gK+ (permeability of K+ channels).
Refractory Period
APD means action potential duration which is marked from the beginning of action potential to the ending of the action potential. For a neuron it is only from 1 to 3 milliseconds. In the case of the Skeletal muscle the duration is 2 to 4 milliseconds. Myocardium 200- 400 milliseconds.Smooth muscles are variable (10-50 milliseconds).
In its wake, the action potential leaves the Na+ channels inactivated and K+ channels activated for a brief time. These transitory changes make it harder for the axon to produce subsequent action potentials during this interval, which is called the refractory period. Two types of refractory periods.
| Absolute refractory period | Relative refractory period |
| It is the sum of the total depolarization period and 1/3rd of the repolarization period. | The last 2/3rd of the repolarization period is nothing but the relative refractory period. |
| A second action potential is not possible in this area. | A second action potential is possible but with the higher Strength of stimulus only. |
| Most of the Na+.V are at the inactive state or open State. | Most of the Na+.V moved from the inactive to the closed stage. |
Physiology Related Articles:
Different Stages of Voltage-Gated Sodium Channel
For instance, in a membrane there is a voltage-gated sodium channel; one of the gates of the sodium channel would be called the M gate. The M Gate is blocking the inner mouth of the channel and the other gate is known as the H gate. So, when the membrane is at resting voltage means the interior of the membrane will be negatively charged and the exterior will be positively charged. This will be known as the closed state of the channel. When the inner voltage of this membrane turns positive or depolarisation occurs this positivity will cause repulsion of the M gate positive charge because of this the gate will be shifted away from the mouth of the channel. This results in opening of the channel and huge amounts of Na+ entering the cell. This stage is called the open stage of voltage gated Na+ channel. If the membrane voltage is maintained at a depolarization State for a long period of time, it will cause the H Gate to block the mouth of the sodium channel. Although the M gate is still away from the channel the H gate will still block the mouth. Na+ will not be able to enter the cell. This stage is known as the inactive stage of the voltage gated Na+ channel.
In the case of hyperkalemia (where Potassium is very high in ECF), a new equilibrium will form. In that equilibrium, the movement of Potassium from inside to outside would be lesser because the charge of the outside Potassium is very high. Ideally, this potassium should move out of the cell but in this case, the potassium stays inside. If there is persistent depolarization the voltage-gated sodium channel will be inactive which will not generate any action potential. And no action potential means no systole in the heart and will result in Diastole.
Hypokalaemia: In the case of hypokalaemia the cell will lose more potassium due to leaky channels, which will raise the negative charge of the cell which results in Hyperpolarization of RMP Which means an increase in RMP. Hypokalemia is also a problem for active myocardium cells. Na+-K+ ATPase will also be inhibited.
In hypocalcemia, when the blood calcium is normal or when the blood calcium is high then only the calcium will remain attached to the extracellular mouth of this voltage-gated sodium channel. And if the blood calcium is low now the main target is to maintain the blood calcium not to occupy the voltage-gated sodium channels that were attached at the mouth of the channel will now detach and because of the detachment the mouth of the sodium channel will open up and the sodium entry will become easier. Due to the increase of the sodium entry, it can be said that the sensitivity of the Na+.V will increase which results in action potential generation leading to an increase in the excitability of the Neuronal membrane.
Nerve Fibre Classification
Classification of Nerve Fibres ;
Depending on the conduction velocity and the diameter and some functional properties, the classification of nerve fibres has been made. This classification of nerve fibres of the body can be seen in the table above. This classification was first done by two scientists, Erlanger and Gasser.
They may be classified as:
- A type of fibre,
- B type of fibre, and
- C type of fibre.
The A fibres have again subdivided into A Alpha, A Beta, A Gamma, and A Delta.
Type B and C do not have any sub-divisions.
The A fibre and B fibres are myelinated fibres. The C fibres are unmyelinated fibres. It was earlier discussed that myelinated fibres will have more conduction velocity. Thus, A and B fibres will have more conduction velocity as compared to the conduction velocity of the C fibres, which are unmyelinated in type. When this classification was proposed by Erlanger and Gasser, later on, two other scientists Lloyd and Hunt proposed another type of classification. The Lloyd and Hunt classification does not differ much. They have introduced numbers instead of the alphabet. They said that A Alpha type of fibres will be known as 1a and 1b. A Beta type of fibre will be known as type II fibre. A Delta type of fibre will be known as type III type of fibre. Type C fibre will be known as type IV fibre. Type B has not been included in their classification. A Gamma has also not been included in the classification.
The function of the Fibres
Now the functions of these fibres will be studied as also the location of these fibres. The Alpha fibres are present in all the proprioceptor fibres in our body and the somatic motor. Proprioception is the position of the joint sense. When one who is closing their eyes is to bring their arm into the same position as the other arm, they can do it. The position of the joint whether in flex or an extended position can easily be understood with the help of this sense. This sense is carried on through the help of some receptors known as proprioceptor receptors. A Beta type of fibre is present in all the touch and pressure fibres. A Gamma fibres are efferent to muscle spindles. A Delta is for pain and temperature. The temperature here means Cold temperature. The C fibre is also for temperature and pain, but here the temperature is warm. Pain is carried by A Delta fibre and also by the C fibres. But the A Delta is myelinated, and the C fibre is myelinated. The pain that is carried by the A delta fibre, has a higher conduction velocity and is therefore known as fast pain. The pain that is carried by the C fibre is known as slow pain. In between the B fibre is the autonomic fibre. It is the preganglionic autonomic fibre. The postganglionic sympathetic fibre is the C fibre.
Also read: Last 5 Year PYQs in Microbiology for INI-CET
The Conduction Velocity of the Nerve Fibres
There is movement from A fibre to C fibre, because there is movement from myelinated to unmyelinated type of fibre, and because the diameter of the fibre is also decreasing from fibre A to fibre C, conduction velocity also decreases. The fastest conducting fibre in our body is the A Alpha variety fibre. This fibre has a conduction velocity of 70 to 120 m/sec. This is proprioception fibre and motor neuron fibre. Proprioception is very important for the maintenance of body balance and posture and that is why the conduction velocity is so huge. Therefore, whenever the posture is disturbed, it will be corrected within fractions of seconds. Similarly, when discussing the touch and pressure fibres the conduction velocity is 30 to 70 m/sec. This gradually decreases to 15 to 30, then to 12 to 30. Fast pain conduction velocity is 12 to 30 m/sec and slow pain is 0.5 to 2 m/sec.
Diameter of the Nerve Fibres
If the conduction velocity is divided by 6 the diameter of the nerve fibre will be achieved. This is just for quick reference. This is not a formula. However, for type C fibre the diameter is almost the same, that is 0.5 to 2. Whatever the conduction velocity is, the same is the diameter in this case.
Compound Action Potential
If an action potential is recorded from the surface of a nerve fibre, the name of such action potential is compound action potential. The kind of nerve fibre that is being dealt with in this case is mixed nerve fibre. Mixed nerve fibres mean that this will contain different types of nerve fibres. For example, in the diagram, it can be seen that this contains A Alpha, A Beta, B, and C types of nerve fibres. If the stimulation is given on the mixed nerve, marked as the point of stimulation in the diagram. The action potential is also recorded from the surface of the neuron, which is marked with recording electrodes. As can be seen from the diagram both electrodes have been placed on the surface of the nerve fibre. This action potential has to be differentiated from the action potential that was discussed earlier which was recorded from a single neuron.
This type of action potential has been discussed in the previous chapter. It is known as a monophasic action potential. This monophasic action potential has been recorded from a single neuron. For this purpose, one electrode has to be inside the cell and one electrode has to be on the surface of the cell.
Now the monophasic action potential will be compared with the compound action potential. In the compound action potential recording, both electrodes have been placed on the surface of the mixed nerve. When stimulation is given on the mixed nerve surface, the nerve fibres will be activated.
Suppose the A fibre, B fibre, and C fibre are activated. Then they will conduct the action potential towards the recording electrode. Now, during the recording of the action potential, the conduction velocity of different nerve fibres has already been discussed. So, the action potential that is being targeted by the A Alpha fibre has a huge and high conduction velocity.
The action potential that is generated by the A Alpha fibre can be seen in the pink coloured curve beside the diagram. The A Beta fibre is also carrying action potential but the speed is slower than that of the A Alpha fibre. The action potential will reach the recording electrode a little later and will form an action potential similar to the red-coloured curve. The B fibre conducting the action potential will be slower than the A Beta fibre. The action potential generated here will appear even later; it is depicted by the white-coloured curve. The C fibre is also conducting the action potential it will appear at the end as depicted by the green coloured curve.
All of these action potentials will merge with one another, and an action potential with multiple peaks will be achieved. This is a biphasic action potential. The major part of this action potential will also go below the membrane voltage. This action potential is known as a biphasic action potential, Multiple peaks of the action potential are present here, It does not follow the "All or none law".
If a higher strength of stimulation is given here and a greater number of A Alpha fibres are activated. Suppose initially 50 A Alpha fibres were activated. Now when the strength of stimulation is increased 100 A Alpha fibres are activated. This will create more voltage because simultaneously many fibres are active. This will cause the amplitude of action potential to be increased. Be all or none law means that the amplitude of action potential is fixed. Therefore, the compound action potential is not following the “All or none law”.
| Compound Action Potential | Monophasic Action Potential |
| This happens in more than one phase and hence is biphasic. Multiple peaks are here due to the activation of multiple fibres. This does not follow all or none law. | This happens in one phase. This has only one peak. |
Also read: Last 5 Years PYQs in Physiology for NEET PG
Stimulus Parameters
Whenever discussing compound action potential, another property has to be kept in mind which is the stimulus parameter. When the stimulus is given on the surface of the neuron, how much voltage is given? That is the amplitude of the stimulus. The second question lies that the stimulus is being given for how long? That is the duration of the stimulus.
A diagram is drawn: the y-axis denotes the amplitude of the stimulus and the x-axis denotes the duration of the stimulus. Amplitude and duration are the two main parameters of the stimulus. Suppose there is a stimulation machine and with the help of this machine stimulation is being delivered to a neuron. In that case, parameter one would be the amplitude of the stimulus and the duration of the stimulus. The third parameter is at what interval such simulation is being given. The interval between the two stimuli will determine the frequency of the stimulation. So, amplitude, duration, and frequency.
Depending on the amplitude, frequency, and duration, three very important terms have to be discussed.
- Rheobase.
- Utilization time.
- Chronaxie.
- Rheobase:
This is the threshold voltage. Suppose a tissue is taken, for example, a neuron or skeleton muscle. If one action potential is to be generated on the tissue by the means of stimulation. Suppose 1 millivolt of stimulus is given and no action potential is received. The strength of the stimulation will then be increased to 2 millivolts. Again, no action potential is seen. Now when 3 millivolts of stimulus are given, an action potential is seen for the first time. After this, if 4 or 5 millivolts of stimulus is given action potential will be generated.
The minimum strength of stimulus voltage that is capable of producing action potential on the tissue is the Rheobase voltage or the threshold voltage. Since 3 millivolts of voltage are enough to generate an action potential, 4 or 5 millivolts of voltage is known as the suprathreshold stimulus for the tissue. Rheobase voltage means it is the minimum strength of stimulus voltage that produces an action potential.
Utilization time
Suppose 3 millivolts of stimulus are being given to the tissue, the voltage is equal to the Rheobase voltage. 3 millivolts is being given for a time of 1 millisecond. No action potential is seen on the tissue. The same Rheobase voltage is given for 2 milliseconds. No action potential is generated on the same tissue.
If 3 millivolts of Rheobase voltage is given for 2.5 milliseconds, suddenly action potential is generated.
3 millivolts Rheobase voltage with a minimum duration of 2.5 milliseconds is sufficient to produce an action potential. This is known as utilization time.
Utilization time is a task in that you are a region of the stimulus. It is the minimum duration of Rheobase voltage that is required to generate an action potential.
Chronaxie
In the previous example, the Rheobase voltage was 3 millivolts, which will now be doubled for this example. 6 millivolts are given for a duration of 1 millisecond on the same tissue, but no action potential is generated. Again 6 millivolts of voltage are being given for 2 milliseconds. Suddenly, an action potential is generated. When stimulus amplitude is increased the duration becomes lesser. Thus, an action potential is generated with a lesser duration of the voltage.
This duration which is required to double the Rheobase voltage to generate an action potential is known as the chronaxie.Chronaxie can be defined as a minimum duration of double the Rheobase voltage that is required to generate an action potential.
Also read: NEET PG Previous Year Question Papers of Last 7 Years
Excitability of the Tissue
With the help of these three parameters one can access the excitability of the tissue. Suppose three tissues are taken, one is neuronal, one is skeletal, and one is myocardial. In order to know which is the more excitable tissue one has to check the chronaxie of these tissues, the utilization time of these tissues, and the Rheobase time of these tissues. The less the chronaxie, the less the utilization time, the less the Rheobase, and the more the excitability of the tissue.
- The most excitable tissue in our body is the neuron, which is followed by skeletal muscles, which are again followed by the myocardium. That is again followed by the smooth muscles.
- This proves that the neuron is the most excitable tissue. The A fibre is the most excitable followed by the B fibre followed by the C fibre. Because the neuron is the most excitable tissue in the body it will have the least chronaxie, the least utilization time and the least Rheobase. However, the most important parameter between these three is chronaxie. That is why when excitability is being checked the chronaxie parameter is also checked.
- Sometimes these three parameters are also plotted in a graphical representation. The graph in which this chronaxie, Rheobase, and utilization time are plotted is known as the strength-duration curve.
Strength-Duration Curve
In this curve, the x-axis represents the time, which is the duration of this stimulus. The Y axis represents the strength, which is the voltage of the stimulus. Suppose a strength-duration curve has been plotted from a neuron. The curve will look like it does in the diagram above. The meaning of this curve is that if any point is taken on the curve, if a perpendicular is drawn on the y-axis, and if a perpendicular is drawn on the x-axis, these points are named x1 and y1 respectively. If a stimulus is given in which y1 is the amplitude and x1 is the duration, this stimulus is capable of generating action potential on the neuron.
So, when any point is taken on the curve, and the strength and duration of that point are found, then it can be concluded that much strength and that much duration is capable of producing one action potential on the tissue. So, the strength-duration curve means one is plotting the stimulus. All these stimulus parameters are capable of generating one action potential in the tissue.
Injury to Nerve Fibers
There is a physiological classification called Sunderland’s classification based on severity of injury.
- There is a parallel classification called Sedon’s classification from the point of view of surgical treatment.
Sunderland’s Classification
- It is 5 degrees of injury.
- 1° injury means mild pressure on the nerve trunk for some time. (Hypoxia)
- 2° → Severe/Sustained pressure

- 3° → Real transaction
- Involves axonal transaction
- 4° → Nerve fascicle disrupted
- 5° → Nerve trunk transected.
- If we talk about big nerves (nerve transactions), those nerves contain bundles of nerve fibres. The bundles are called fascicles & each fascicle is made up of a few axons or nerve fibres.
- 3° injury means cutting of just one axon.
- 4° injury would be cutting off the entire nerve fascicle.
- 5° injury would be nerve trunk transaction which means the entire nerve trunk has been cut through & through.
Seddon’s classification
- In Seddon’s classification, the pressure related injuries are called Neuropraxia.
- One axon cut is Axonotmesis.
- More than one axon cut, or a bundle cut is called Neurotmesis.
- What happens after the injury?

- These changes described happen after 5° injury when the nerve trunk is transacted.
- In 5° injury the nerve trunks are cut in two pieces. These nerves of neurons have nerve cell bodies.
- When injury happens, two segments are formed.
- The segment towards the nerve cell body is called Proximal segment.
- The segment away from the cell body is called Distal segment. The degenerated changes will be seen in the Distal segment.
Changes

- 24 – 48 hours after the transaction, there is appearance of Chromatolysis.
- Chromatolysis means dissolution of the Nissl granules. The nerve cells imbibe water.
- The nucleus is pushed to the periphery .
- All these changes are collectively called chromatolysis.
- It is thought to be a preparation for healing.
- Up to 3 days, the distal stump is functional. After all, it is a nerve fibre, the job of nerve fibre is to conduct the impulses, so for 3 days it will carry the impulse but after 3 days, its gone & there is no sensation. It will lose its function.
- By 6th day after the injury, there will be axonal degeneration. Axis cylinder breaks. By the 10th day, there will be myelin degeneration. So there are just some oil droplets in an empty tube. It is because the Schwann cell & its membrane is rich in lipids.
Also read: INI-CET Previous Year Question Papers
Sarcomere
Structure of the Sarcomere
- Inside the muscles, there are muscle fibres and within each muscle fibre, there is the sarcomere. Sarcomere is the distance between the two successive z-lines.
- At regular distances, there are Z lines (zigzag).
- Either side of Z-lines, there are thin filaments that arise. Among the thin filaments, there are thick filaments.

Length of the Sarcomere
- The normal resting length of the sarcomere is 2 microns. It is called L initial or L resting which is 2.0μ.
Starling’s Law
- Greater the initial length of the muscle, stronger the contraction within physiological limits.
- If we stretch the muscle in its resting condition, sarcomere length will also increase.
- 2.1 μ (strong contracting)
- L0 – 2.2μ (strongest contraction)
- If we stretch the sarcomere further, beyond this length the contraction will become weak, thick filament overlap will be lost. So from this length, contraction will become weak
- Lmax = 3.65 μ−3.7 μ
- This is the maximum resting initial length of the sarcomere & it will produce hardly any contraction, almost no contraction.
- If we take the cross – sectional view of the Z line, it is a hexagon. From all its 6 angles arises one thin filament.
- In these thin filaments are interspersed thick filaments. Each thick filament is surrounded by 6 thin filaments. Each of the thin filaments is surrounded by 3 thick filaments.
- Therefore, the ratio of thin filaments in a skeletal muscle sarcomere is 2:1.

Muscle proteins
1. Dystrophin
- It starts from the thin filament & it reaches the sarcolemma. In the sarcolemma there is a complex of six glycoproteins, 2 subunits of dystroglycan and 4 subunits of sarcoglycans.
- The β– Lema is attached to an extra cellular matrix protein which is called Laminin.
The function of dystrophin
- Thin & thick filaments will interact & they will generate tension in the muscle. Transferring that tension to the exterior of fibre & to the tendon of the muscle is the job of dystrophin.
- The dystrophin gene is the largest gene.
- Defect of dystrophin leads to Duchenne’s muscular dystrophy
2. Titin

- Largest known human protein.
- It starts from the Z- line & enters the thick filament.
- Its function is the alignment of thick filament in the sarcomere.
- The genetic defect of Titin results in: Limb girdle muscular dystrophy
Also read: Solved Numericals In Pharmacology NEET PG
Important Points to Remember
- At Lo muscle contraction is strongest due to Frank Starling's law (greater the initial length - strongest is the contraction) beyond this length (Lo) the sarcomere will become weak and strength of contraction decreases.
- Goldman's constant field/ Goldman's Hodgkin Katz Equation Considers 2 Factors
- Concentration gradient of individual ions
- Membrane permeability
- Hair cells are surrounded by Endolymph which is having excess of K+ Due to which its potential becomes + 80 mv, known as endocochlear or endolymphatic potential.
Important Solved Questions
Q. In a skeletal muscle sarcomere, each thin filament is surrounded by how many thick filaments?
A. 3
B. 4
C. 6
D. 8
Ans: 3
Q. Which of the following cells has the highest transmembrane electrical gradient?
Ans. Hair cell
- If skeletal muscle is – 90 mV on the inside and exterior is taken to be 0, it is a relative voltage. So, the trans-membrane electrical gradient will be 90 mV.
- There is one cell in the body where transmembrane voltage gradient of 150 mV & that is the highest for any cell in the body. It is the hair cell in the labyrinth. It is the highest because inside the membrane is –70 mV but outside is not zero. Hair cell in the labyrinth is surrounded by a very exceptional ECF called Endolymph. Endolymph has got excess K+ ions.
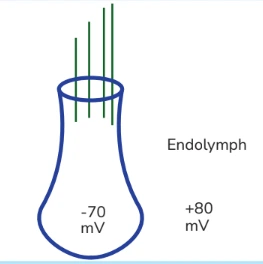
- It is an ECF & K+ should be low but here there are excess K+ ions over & above the ECF. So its voltage is not zero but something above zero because of these +ve charges. It has got +80 mV. It is called Endocochlear potential or Endolymphatic potential.
- 150 mV is the highest electrical gradient in the body which is for the hair cell. It makes it the most excitable cell in the body.
Q. There is a cation. Its ECF concentration is 100 & ICF is 10. Calculate its equilibrium potential.
Ans. It is a cation & because of its concentration gradient, it will move from outside to inside initially. It will carry the positive charges to the inside & then eventually it will reach equilibrium. When it reaches equilibrium, the charge on the membrane will be +. After putting the values in the equation, equilibrium potential for this cation will be +61 mV.
Q. If there was an anion & its concentrations are 100 on the outside & 10 on the inside. Calculate the equilibrium potential for this anion.
Ans. – 61 mV
Because all other things will remain the same. Since this is an anion & by its initial concentration, it will go from outside to inside & carry the negative charges to the inside. So, when it reaches equilibrium, charge on the membrane will be –ve.
Nerve
- RMP of a nerve = – 70mV
- Equilibrium potential for Na+ = + 60 mV
- Equilibrium potential for K+ = –90 mV
- Equilibrium potential for Cl- = -70 mV
- Na+ concentration will remain the same in the extra cellular fluid but in the intra cellular, the nerve will have a different value for Na+ and muscle will have a different volume of ICF sodium.
- RMP of nerve is –70 mV & Equilibrium potential for Cl- is also –70 mV which means when the nerve is at RMP:
- EP for Ca++ = +129 mV (highest)
- Ca++ is a divalent ion. It brings double +ve charge
- EP for H+ = – 23 mV
- EP for HCO3– = –24 mV
Q. First change seen in the Wallerian degeneration is:
Ans. Axonal degeneration
Q. First change seen after nerve trunk transaction is:
Ans. Chromatolysis
- Regeneration & repair starts by 15th day after the injury. Two changes happen almost simultaneously. Sprouting from the proximal segment & Schwann cell proliferation from the Distal segment.
- Repair is completed by the 80th day.
- Axons never reach its original diameter. It only reaches 80% of its original diameter.
Also read: Genetics Questions Solved for NEET PG Biochemistry Preparation
This is everything that you need to know about nerve muscle physiology for your physiology preparation... For more interesting and informative blog posts like this download the PrepLadder App and keep reading our blog!


PrepLadder Medical
Get access to all the essential resources required to ace your medical exam Preparation. Stay updated with the latest news and developments in the medical exam, improve your Medical Exam preparation, and turn your dreams into a reality!
Top searching words
The most popular search terms used by aspirants
- NEET PG Physiology
- NEET PG Strategy
PrepLadder Version X for NEET PG
Avail 24-Hr Free Trial